Selektion
1. Was ist Selektion?
Die Selektion ist ein Vorgang in der Natur und bezeichnet die natürliche Auslese durch die äußeren Lebensbedingungen, sprich der Umwelt. Den Vorgang der Selektion gibt es schon seitdem es Lebenswesen gibt, vielleicht sogar schon davor. Vorraussetzung für Selektion ist eine genetische Variabilität innerhalb der Art, die wiederum notwendigerweise Mutation voraussetzt. Da Mutationen schon seit dem Entstehen erster Lebewesen auf der Erde vorkamen, kann man davon ausgehen, dass Selektion schon seit dieser Zeit stattgefunden hat und die Evolution erst ermöglicht hat.
Zur Erläuterung meiner Vermutung, dass es vielleicht schon vor den ersten Lebewesen Selektion gab, also ohne genetische Variabilität, möchte ich folgendes sagen. Ich gehe davon aus, dass es vor dem, was wir heute als Lebewesen bezeichnen, schon andere selbstreplizierende Moleküle gab. So etwas wird als präbiotischer Replikator bezeichnet. Die Selektion besteht nun darin, dass ein zufällig gebildetes Molekül den Anforderungen der Umgebung und der Selbstreplikation gerecht wird. Zufällige "Fehler" im Aufbau dieser Strukturen können zu deren Verbesserung beitragen, was ihnen bei der Replikation einen Vorteil verschafft. Die Selektion wirkt also auf zufällig entstandene Moleküle ein, die durch eine Mutation zufällig weiter verbessert werden können. Diese Selektion ist also analog mit der bei Lebewesen heutiger Form zu sehen.
1.1 Selektionsprinzip nach Darwin
Das Selektionsprinzip ist eine der Grundannahmen von Charles Robert Darwins1 Evolutionstheorie, die größtenteils auf den Erkenntnissen beruht, die er während einer fünfjährigen Forschungsreise auf dem Vermessungsschiff Beagle gewann. Dieses Prinzip gründet sich auf folgenden Begründungen und Erfahrungen:
-
Die durch die Umwelt vorgegebenen Ressourcen, die für ein Überleben der Art sorgen, sind für jede Population knapp.
-
Einzelne Individuen einer solchen Population, die über Eigenschaften verfügen, durch die sie die knappen Ressourcen besser nutzen können haben einen Überlebensvorteil, weil sie einen Vermehrungsvorteil haben.
-
Dies geschieht durch einen immerwährenden Kampf um die Ressourcen, einen Kampf ums Dasein (struggle for life), der zum Überleben der Lebewesen führt, die am besten an die derzeitige Umwelt angepasst sind (survival of the fittest).
Dieses Selektionsprinzip gilt in seinen Grundzügen noch heute, nur dass die Erklärungen des beobachteten Phänomens heutzutage auch quantitativen Charakter angenommen haben, d.h. es lassen sich auf mathematischem Wege präzise Vorhersagen über die Entwicklung einer bestimmten Population treffen. Dies ist aber erst möglich geworden, als man die Vererbungstheorie hinreichend weit entwickelt hatte.
1.2 Grundlegende Begriffe der Selektion
Dieser Abschnitt ist eine Art Glossar in dem eine Erklärung der wichtigsten Begriffe der Selektion geboten wird. Wer bereits ein Grundlagenwissen im Bereich der Selektion hat kann diesen Abschnitt überspringen. Begriffe, die im Glossar erklärt sind werden im weiteren Verlauf nicht noch einmal erklärt, es sei denn sie werden im Text weiterentwickelt.
Selektionsfaktor
Umwelteinfluss, der unterschiedliche Fortpflanzungsraten verschiedener Phänotypen bewirkt.
Selektionsdruck
Einwirkung der Selektionsfaktoren auf eine Population. Bewirkt eine Veränderung des Genpools.
Stabilisierende Selektion
Transformierende Selektion
Disruptive Selektion
Bekannt als die drei Wirkungsweisen der Selektion. Geben an in wiefern sich der Genpool und damit die Häufigkeiten der Phänotypen verändert.
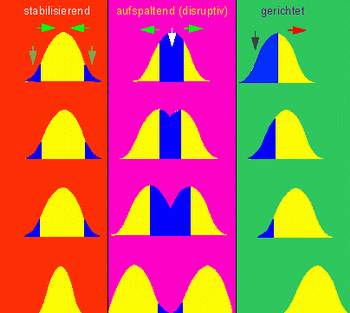
Bei der stabilisierenden Selektion werden, Phänotypen begünstigt, die in der Mitte der derzeitigen Phänotypenvariabilität liegen. Diese wird dadurch reduziert und führt zu einer Erhaltung des Ist-Zustands. (Abb. 2 links)
Bei der transformierenden Selektion (gerichtete Selektion) verschiebt sich die Mitte der derzeitigen Phänotypenvariabilität in eine gewisse Richtung. (Abb. 2 Mitte)
Bei der disruptiven Selektion, spaltet sich die Phänotypenhäufigkeit, so dass beide Extrema begünstigt werden. Man könnte es auch als transformierende Selektion in beide Richtungen ansehen. (Abb. 2 rechts)
Fitness
Größe für den Fortpflanzungserfolg eines Individuums in der nächstfolgenden Generation. Dieses Maß ist Ausdruck der gesamten Lebensleistung des Individuums unter den Bedingungen der natürlichen Selektion.
Genetische Variabilität
Innerhalb einer Art auftretende voneinander abweichende Merkmalsprägungen (Varianten) in der Erbinformation. Die genetische Variabilität spiegelt die gesamte Bandbreite an genetischen Möglichkeiten wieder, die ein einzelnes Gen in einer Art annehmen kann. Somit ist sie für die Anpassungsfähigkeit von Populationen entscheidend.
Genpool
Gesamtheit der Gene in einer Population, wobei die genetische Variabilität maßgebend für die Heterogenität des Genpools ist.
1.3 Wie wirkt die Selektion?
Die Selektion wirkt auf sehr mannigfaltige Weise. Sie ist ein natürlicher Vorgang der nicht zielgerichtet verläuft, sondern einfach eine Tatsache darstellt, die da lautet: "Das durch seine Gene am besten angepasste Individuum hat die größte Fitness und somit die größte Chance zu überleben, wodurch auch sein Fortpflanzungserfolg steigt, was wiederum Auswirkungen auf den Genpool hat."
Diese allgemeine Aussage kann nun auf spezielle Fälle bezogen werden, bei denen man nur ein paar Merkmale - vielleicht sogar nur ein einzelnes - betrachtet, um Aussagen über den Nutzen verschiedener Erscheinungsformen zu treffen. In diesem Zusammenhang ist es wichtig zu erkennen, dass die Selektion nur auf den Phänotyp eines Individuums einwirkt, nicht aber auf den Genotyp. Mit anderen Worten, entlehnt aus der Mendelschen Genetik, gesprochen: "Bei einem dominant-rezessiven Erbgang hat ein AA-Individum hat die gleiche Fitness wie ein Aa-Individum." Diese Tatsache ist von außerordentlicher Wichtigkeit, denn auf diese Weise bleibt die genetische Variabilität erhalten, die nötig ist bei einer veränderten Umwelt den Fortbestand der Art zu sichern. Da ein Aa-Individuum nämlich immer noch das vermeidlich schlechte rezessive a-Allel enthält, es aber die selbe Überlebenschance hat wie ein AA-Individuum, entstehen in jeder neuen Generation immer wieder ein paar AA-Individuen, die zum jetzigen Zeitpunkt im Nachteil sein mögen, der sich aber auch schnell zu einem Vorteil entwickeln kann, wie wir später noch sehen werden. Natürlich ist dieser Fall grob vereinfacht, weil nur ein Gen für die Ausbildung eines bestimmten Merkmals verantwortlich ist und ein dominant rezessiver Erbgang vorausgesetzt wurde. Oft ist es so, dass bis zu hundert Gene ein bestimmtes Merkmal beeinflussen. In solchen Fällen ist es schwerer nachzuvollziehen, was von Vorteil und was von Nachteil ist. Die Komplexität wird noch dadurch gesteigert, dass Gene auch auf mehrere Merkmale Einfluss nehmen können. In solchen Fällen helfen nur noch komplexe mathematische Modelle, um die Entwicklung einer Population vorhersagen zu können.
2. Selektionsfaktoren
2.1 Abiotische Selektionsfaktoren
Zu den abiotischen Selektionsfaktoren zählen all diejenigen Selektionsfaktoren, die durch unbelebte Materie hervorgerufen werden. Zu ihnen zählen Sonneneinstrahlung, Wind, Salzgehalt von Wasser, Temperatur, Naturkatastrophen, Sauerstoffgehalt von Luft oder Wasser. Auch hier kann man keine eindeutige Grenze ziehen. So kann der Sauerstoffgehalt von Wasser auch auf eine vom Menschen verursachte Eutrophierung des Gewässers hinweisen. Auch kann man sich darüber Streiten, ob das Fallbeispiel des Birkenspanners, wirklich in den Bereich der abiotischen Selektionsfaktoren fällt. Denn, wie wir noch sehen werden, hat der Mensch an dieser Entwicklung einen nicht unerheblichen Anteil. Ich werde im nächsten Abschnitt , warum ich mich dazu entschlossen habe es doch zu den abiotischen zu zählen.
2.1.1 Fallbeispiel Birkenspanner
Der Birkenspanner Bison betularis ist ein Nachtfalter, welcher feinscheckig gemusterte Flügel besitzt. Der Birkenspanner sitzt tagsüber oft an Baumrinden, da er sich von den am Baumstamm lebenden Flechten ernährt. Seine Flügel- und Körperfärbung ermöglicht es ihm tagsüber, vor seinen Fressfeinden, wie bespielsweise Singdrosseln, Rotkehlchen und Kohlmeisen, weitesgehend unentdeckt zu bleiben, da er vor dem Hintergrund der hellen Baumflechten schwer zu erkennen ist.
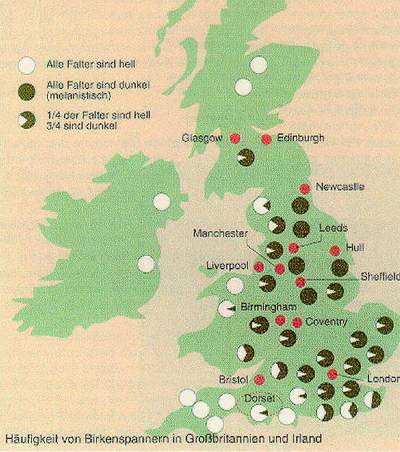
Durch eine Mutation entstanden zufällig aber auch immer einige dunkle Exemplare. Diese Tiere haben eine gesteigerte Produktion des Farbstoffs Melanin. Dies ist auf ein dominant wirkendes Melanismusallel zurückzuführen. Somit stellt der Melanismus das Gegenstück zum Albinismus dar, nur das sich dieser rezessiv vererbt. Normalerweise stellt der Melanismus einen Nachteil dar, da der Flechtenhintergrund an den Baumstämmen, auf dem die Falter sitzen, in aller Regel hell ist wie folgende Abbildung zeigt.

Nun müsste die dunkle Form dadurch einem starken Selektionsdruck seitens der Fressfeinde ausgesetzt sein wodurch sie, auch wenn sich Melanismus dominant vererbt, nicht sehr verbreitet sein dürften. Wie kommt es dann, dass heutzutage in allen Industriestädten Englands, die Population fast ausschließlich aus dunklen Individuen besteht?
Nun der Selektionsdruck auf die hellen Individuen müsste wesentlich größer sein als der auf die dunklen, aber warum ist das so?
Die Ursache dafür ist in der starken Luftverschmutzung zu sehen, die durch die Fabrikanlagen in den englischen Städten verursacht wird. Durch die Verschmutzung sterben die Baumflechten ab und die Rinde färbt sich dunkel. Dadurch sind die hellen Falter eine leichte Beute für ihre Fressfeinde, ähnlich wie ein Albinohase praktisch keine Chance hat einem Fuchs oder einem Luchs zu entkommen weil er einfach zu auffällig ist und keine Tarnung hat.

Nun war die helle Form des Birkenspanners einem starken Selektionsdruck ausgesetzt. Dies führte zu einer Veränderung des Genpools. Die Selektion hatte transformierende Wirkung. Aus denen in 1.3 genannten Gründen treten aber immer auch helle Nachfahren dunkler Eltern auf, so dass der Genpool irgendwann keine Änderung mehr erfährt. Ist die Transformation abgeschlossen, hat die Selektion stabilisierende Wirkung.
Auch in den weniger industrialisierten Teilen der Britischen Inseln, ist die helle Variante noch vorherrschend, wie folgende Abbildung zeigt. Der Osten Englands hat mehr dunkle Formen, da der Westwind vorherrschend ist.
Nun zur Begründung, warum dieses Fallbeispiel unter den abiotischen Selektionsfaktoren auftaucht. Schließlich ist die Luftverschmutzung doch eine vom Menschen verursachte Wirkung, also ein biotischer Faktor.
So könnte man sicherlich argumentieren, doch der eigentliche Selektionsfaktor ist doch die Tarnung, also welche Form ist gegenüber ihren Fressfeinden am besten getarnt. Wer am besten getarnt ist entscheidet die Umgebung. Die Baumstämme sind dunkel, weil die Verschmutzung die Flechten abtötet. Die Verschmutzung hätte aber genausogut durch einen Vulkanausbruch oder eine ähnlich Naturkatastrophe herbeigeführt werden können. Den Mensch als biotischen Faktor anzusehen birgt eine große Gefahr in sich, da dies eine Rechtfertigung für das gesamte menschliche Handeln darstellen würde. Die Natur hat uns geschaffen, also ist sie selber schuld, wenn wir sie verseuchen. Der Unterschied zwischen dem Menschen und allen anderen auf der Erde vorkommenden Lebewesen ist, dass der Mensch die Folgen seines Handelns übersehen kann. Er kann sich nicht aus der Verantwortung stehlen. Eigentlich bildet der Mensch eine eigene Gruppe von Selektionsfaktoren. So hätte Anfang des zwanzigsten Jahrhunderts ein Bär enorme Vorteile gehabt, dessen Fell ungeeignet zur Pelzherstellung wäre. Aufgrund dieser Gefahr und der möglichen Ersetzunng des Menschen durch einen "wirklichen abiotischen" Faktor habe ich mich dazu entschlossen dieses Fallbeispiel unter die abiotischen Selektionsfaktoren zu zählen.
2.1.2 Fallbeispiel Spitze Strandschnecke
Die Spitze Strandschnecke Littoria littorea bewohnt die Spritzwasserzone an Meeresküsten. Auf sie wirken gleich mehrere abiotische Selektionsfaktoren ein.
Zum einen ändert sich der Salzgehalt ihrer Umgebung durch die Gezeiten ständig. Bei Ebbe steigt er, bei Flut sinkt er dann wieder. Auch die Umgebungstemperatur unterliegt enormen Schwankungen, die teilweise durch die Gezeiten verstärkt werden.
In dieser unfreundlichen Umgebung ist die Strandschnecke auf ihr Gehäuse angewiesen. Bei anderen Schneckenarten dient es hauptsächlich dem Schutz vor Fressfeinden, aber die Strandschnecke nutzt das Gehäuse auch, um Feuchtigkeitsgrad, Salzgehalt und Temperatur im inneren des Gehäuses stabil zu halten.
Der Selektionsdruck bewirkt also, dass Individuen, die zufällig besser an diese Bedingungen angepasst sind, besser überleben können, so mehr Nachkommen bekommen und dadurch diese ökologische Nische besetzen. Man kann außerdem davon ausgehen, dass die am besten angepassten Individuen dieser Art am höchsten in der Gezeitenzone siedeln, da sie hier weniger Konkurrenz von Artgenossen haben und so mehr Nahrung finden.
Dieses Beispiel verdeutlicht sehr schön die Wirkungsweise der Selektion und somit auch den Weg den die Evolution beschreitet. Ursprünglich war das Gehäuse der Schnecken nur dazu da Schutz zu bieten, doch unter diesen spezifischen Lebensbedingungen kann es eine Doppelfunktion übernehmen. Was ich damit zum Ausdruck bringen will ist, dass die Evolution nicht alle Merkmale einer Art gleichzeitig über den Haufen werfen kann. Da Mutationen zufällig geschehen, finden immer nur kleine Veränderungen in einem bestimmten Rahmen statt. Die Selektion wirkt also vor allem in der Weise, dass sie Verbesserungen bereits vorhandener artspezifischer Merkmale begünstigt.
2.1.3 Fallbeispiel flugunfähige Insekten
Dem letzten Absatz im vorherigen Abschnitt folgend, sei hier ein weiteres Bespiel für Evolution innerhalb der vorgegebenen Grenzen genannt.
Auf den Kerguelen, einer kleinen Inselgruppe im südlichsten Pazifik leben, viele Insekten, die solch eine Gestalt aufweisen.



Doch warum sind die Flügel zurückgebildet? Dies müsste unter normalen Umständen doch einen Nachteil für die Tiere darstellen, doch offensichtlich wird auf die Insekten ein starker Selektionsdruck ausgeübt, so dass Flugunfähigkeit vorteilhaft ist.
Und tatsächlich herrschen auf den Kerguelen keine "normalen" Wetterbedingungen. Die Kerguelen sind starken Winden ausgesetzt, die oft sogar Orkanstärke erreichen. Fliegende Insekten würden leicht vom Wind aufs offene Meer getrieben werden und dort umkommen. Infolge von Mutationen sind die Flügel also immer weiter verkümmert, bis die Insekten überhaupt nicht mehr fliegen konnten. Die Flügel sind aber immer noch vorhanden, was wiederum auf den Weg der Evolution als Weiterentwicklung bereits vorhandener Merkmale hinweist. Dieses Vorhandensein ohne praktischen Nutzen bezeichnet man als rudimentär.
2.2 Biotische Selektionsfaktoren
Biotische Selektionsfaktoren müssen meist in ihrer Gesamtheit gesehen werden, da Tiere und Pflanzen mannigfaltige Wechselwirkungen untereinander eingehen. Will man ein komplettes Nahrungsnetz für den Braunbären erstellen, so hat man einiges vor sich. Die Getrenntbetrachtung einzelner Faktoren macht deshalb nur bei sehr ausgewählten Fällen Sinn.
Dieser Abschnitt ist in zwei große Teilbereiche unterteilt. Der erste befasst sich mit der sexuellen Selektion, der zweite mit der Verbindung zwischen der Sichelzellanämie und Malaria.
2.2.1 Sexuelle Selektion
Die sexuelle Selektion war schon für Darwin ein wichtiger Stützpfeiler seiner Evolutionstheorie. Das unterschiedliche Aussehen der Geschlechter, der sog. Sexualdimorphismus, brachte ihn zu der Annahme, dass es zwei verschiedene Arten der sexuellen Selektion gibt. Zum einen der Kampf der Männchen um die Weibchen und um Reviere zur Versorgung seiner Weibchen (intrasexuelle Selektion). Zum anderen die Wahl des Paarungspartners durch das Weibchen (intersexuelle Selektion).
2.2.1.1 Fallbeispiel Rothirsch
Der Rothirsch Cervus elaphus, ist ein Paradebeispiel für die intrasexuelle Selektion. Alljährlich zur sog. Brunftzeit im Oktober tragen die männlichen Tieren auf Brunftplätzen Kämpfe aus, um so eine kleine Weibchengruppe zu erkämpfen.
Die Konfontationen können aus Röhrduellen, Parallelgehen,einem Kampf oder einer Kombination dieser Formen bestehen. Dabei spielt das Geweih eine entscheidende Rolle. Es ist Symbol der Stärke des Hirsches. Die Geweihgröße ist abhängig von Körpergröße und Gewicht des Hirsches. Kommt es zu einer Annäherung zweier Hirsche so wird durch das Röhren und das Parallelgehen die Kampfkraft des Gegners eingeschätzt. Fühlt ein Hirsch sich unterlegen, so kann er jerderzeit aufgeben und das Weite suchen. Folgende Abbildung zeigt die Verteilung der verschiedenen Entwicklungen einer solchen Konfrontation.
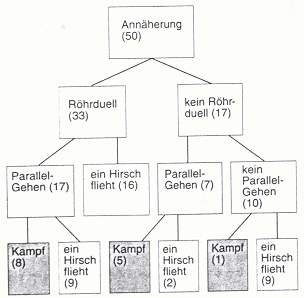
Interessant ist, dass es nur in 28% der Fälle zu einem Kampf kommt. Die Kontrahenten können also sehr gut die Stärke des anderen einschätzen.
Wie schon erwähnt hängt der Paarungserfolg maßgeblich von der Geweihgröße ab. Als Voraussetzung für ein großes Geweih gilt ein hohes Körpergewicht und ein großer Körperwuchs. Die Entwicklung von Körpergewicht und Größe hängt vom Zeitpunkt der Entwöhnung der Muttermilch ab.
Deshalb ernähren ranghohe Hirschkühe ihre Söhne besser als ihre Töchter, da ein ranghoher Hirsch mehr Nachkommen zeugen kann als eine ranghohe Hirschkuh. Rangniedere Hirschkühe versorgen ihre Töchter besser, da eine rangniedere Hirschkuh fast genausoviele Nachkommen bekommen kann, wie eine ranghohe Hirschkuh, ein rangniederer Hirsch aber unter Umständen gar keine Nachkommen zeugen kann. Die Hirschkühe verhalten sich also so, wie sich für sie die größtmögliche Anzahl von Enkeln ergibt; ihr Verhalten ist also adaptiv.
Dies ist ein weiteres Hauptmerkmal der Selektion. Alle Individuen verhalten sich so, dass sie ihre Gene möglichst oft weitergeben. Wir werden im Abschnitt über Verwandtschaft und Selektion noch sehen, dass dies noch etwas präziser formuliert werden muss.
2.2.1.2 Fallbeispiel Rauchschwalbe
Die europäische Rauchschwalbe Hirundo rustica ist ein gutes Beispiel für die intersexuelle Selektion, also die Wahl des Paarungspartners durch das Weibchen. Auffällig ist, dass das Mänchen längere Schwanzfedern hat als das Weibchen. Hier ein Bild einer männlichen Rauchschwalbe.

Die Schwanzfeder des Männchens scheint also ein sekundäres Geschlechtsmerkmal darzustellen. Spielt sie auch beim Paarungserfolg eine Rolle?
Tatsächlich finden Mänchen mit längerer Schwanzfeder schneller ein Weibchen, als Männchen mit kurzer Schwanzfeder und können auf diese Weise eher anfangen zu brüten und so sogar mehrmals brüten. Ihre Vermehrungsrate ist also deutlich höher, als die von Männchen mit kurzer Schwanzfeder. Folglich müssten Rauchschwalben immer längere Schwanzfedern bekommen, da sie einem starken Selektionsdruck ausgesetzt sind, der den Genpool verändern würde. Die durchschnittliche Schwanzfederlänge beträgt 10 cm und eine Änderung ist seit dem die Rauchschwalben wissenschaftlich untersucht werden nicht festzustellen. Es muss also noch einen anderen Selektionsdruck geben, der dem der intersexuellen Selektion entgegengesetzt wirkt.
Tatsächlich hat man festgestellt, dass Männchen mit längeren Schwanzfedern es ungleich schwerer bei der Futtersuche haben. Die lange Schwanzfeder verschlechtert die Flugeigenschaften, so dass Männchen mit überdurchschnittlich langen Schwanzfedern eine niedrigere Lebenserwartung haben. Die beiden Selektionsdrücke bilden sozusagen ein Gleichgewicht heraus, dass nun zufällig bei 10 cm liegt.
Warum suchen sich Weibchen nun also Männchen mit längeren Schwanzfedern aus, wenn sie doch bei der Futtersuche arge Schwierigkeiten bekommen? Die Federlänge sagt auch etwas über das Alter und den Gesundtheitszustand des Männchens aus. Eine lange Schwanzfeder steht also für ein erfahrenes und gesundes Männchen. Kurze Schwanzfedern für ein junges und krankes Männchen.Die Länge der Schwanzfeder ist für die Weibchen eine Art Statusanzeige, denn sie wissen, dass ein erfolgreiches und gesundes Männchen ihre Nachkommen überlebensfähiger macht als ein krankes.
Hier betrachten wir zum ersten Mal zwei Selektionsfaktoren zusammen. Einmal den der intersexuellen Selektion und dann den der Fähigkeit Beute zu machen. Nur die Männchen, die einen guten Kompromiss zwischen den beiden begrenzenden Faktoren gefunden haben, werden sich erfolgreich vermehren können, was zu einer stabilisierenden Selektion führt.
2.2.1.3 Fallbeispiel Bankivahuhn
Das Bankivahuhn Gallus gallus wird oft als Beispiel für eine Kombination aus intra- und intersexueller Selektion aufgeführt. Es findet sowohl ein Kampf der Männchen um die Weibchen statt (Hahnenkampf), als auch die Hennen die Hähne nach spezifischen Merkmalen beurteilen.
Hähne mit großen leuchtenden Augen, großen roten Kämmen und Kehllappen werden von den Hennen bevorzugt (Abb. 9). Diese Merkmale spiegeln wie bei der Rauchschwalbe den Gesundtheitszustand des Hahnes wieder. Gleichsam sind sie auch Ausdruck der Stärke eines Hahnes und dienen zur Einschüchterung von Rivalen.

Da Hähne mit diesen äußeren Merkmalen mehr Nachkommen zeugen können bilden sich die Merkmale immer stärker heraus. Es kann angenommen werden, dass mit diesen phänotypischen Merkmalen auch genotypisch vorteilhafte Anlagen verbunden sind. Die Selektion wirkt in diesem Fall gerichtet in Richtung der oben genannten phänotypischen Merkmale.
2.2.2 Verwandtschaft und Selektion
Zunächst fragt man sich, was Verwandschaft mit Selektion zutun hat. Dazu müssen wir folgende Grundüberlegungen treffen.
Wir sprachen ja bereits vom Begriff der Fitness. Die Fitness ist ein Maß dafür, wie oft ein Individuum seine Gene weitergibt. Also hat ein Individuum mit mehr Nachkommen auch eine höhere Fitness, als ein Individuum mit weniger Nachkommen. Wenn also entscheidend ist, wer seine Gene am häufigsten weitergibt, müsste man dann nicht Individuen einer Art mit ähnlichen Genen zu einer Gruppe zusammenfassen?
William D. Hamilton bezog das erste Mal in solche Überlegungen ein, dass verwandte Individuen eine relativ hohe Übereinstimmung ihrer Gene erreichen. Infolgedessen kann es für Individuen nützlich sein, ihre Verwandten bei der Aufzucht des Nachwuchses zu unterstützen, da diese einen Teil der eigenen Gene enthalten und somit der Nachwuchs auch einen Teil der Gene des helfenden Individuums erhält. Allerdings müsste die Wahrscheinlichkeit einer solchen Hilfe mit der Entferntheit des Verwandten abnehmen. Nach Hamilton kann diese Hilfe nur erfolgen, wenn der Nutzen (N) für den Unterstützten multipliziert mit dem Verwandtschaftsgrad (r) größer ist als die Kosten (K) für den Selbstlosen. Daraus ergibt sich die berühmte Hamilton-Ungleichung:
K < r • N
Insofern müssen wir den Begriff der Fitness erweitern. Ein Individuum kann sein Erbgut nämlich auf zwei Wegen weitergeben, indem es sich selber fortpflanzt (direkte Fitness) und indem es Verwandte bei der Aufzucht unterstützt (indirekte Fitness). Addiert man beide Werte so erhält man die Gesamtfitness eines Individuums.
Mit diesen Vorkenntnissen betrachten wir nun die folgenden zwei Fallbeispiele, das des Buschblauhähers und das der Honigbiene.
2.2.2.1 Fallbeispiel Buschblauhäher
Der Buschblauhäher Aphelocoma coerulescens caerulescens ist ein gutes Beispiel für die Bruthilfe. Er siedelt ausschließlich in Florida. Dort bevorzugt er zum Brüten das Gestrüpp von Eichen. Da der Brutraum sehr begrenzt ist, finden nicht alle Vögel einen Brutplatz. Dann geschieht etwas sehr merkwürdiges. Diese Vögel helfen anderen Artgenossen beim Brüten. Sie beschaffen bis zu 30% des Futters für die Jungen und entlasten auf diese Weise die Eltern. Bis zu zehn Bruthelfer unterstützen ein Brutpaar. Doch wieso sind diese Vögel so selbstlos?

Wir sprachen ja schon über die Hamilton-Ungleichung. Sie ist Teil der von Hamilton entwickelten Theorie der Verwandtenselektion. Diese Theorie hat eine ganz einfache Lösung für dieses Problem. Da der Brutraum begrenzt ist können nicht alle Vögel einen Brutplatz ergattern. Die Vögel, die keinen Brutplatz sichern konnten, haben zwei Möglichkeiten. Entweder sie tun gar nichts oder sie helfen einem anderen Brutpaar.
Man kann sich leicht vorstellen, dass die Gene zum selbstlosen Helfen eigentlich nicht begünstigt werden dürften, da sie ja von den Vögeln, die so selbstlos sind und helfen gar nicht weitergegeben werden können. Man hat aber festgestellt, dass nur mit dem Brutpaar verwandte Vögel diesem auch helfen und zwar der Hamilton-Ungleichung folgend. Diese Gene werden nun natürlich auch begünstigt, da sie im Brutpaar selber auch zum Teil vorhanden sein müssen, da diese mit den Helfern ja verwandt sind. Durch die Helfer steigt nämlich die Überlebenswahrscheinlichkeit der Jungen und auch der Eltern, wodurch ein Selektionsvorteil gegeben ist. Anders ist es bei Genen, die bewirken würden, auch nicht verwandten Vögeln zu helfen. Nicht verwandte Vögel haben höchstwahrscheinlich keine gleichen Gene. Dadurch würden diese "Samariter-Gene" nicht weitergegeben und diese Vögel wären selektiv benachteiligt. Bald würden diese Gene aus dem Genpool verschwinden. Die verwandten Vögel, die gezwungen sind nicht zu brüten, handeln also adaptiv, wenn sie Verwandten bei der Aufzucht helfen, denn auf diese Weise erreichen sie jedenfalls eine teilweise Weitergabe ihrer Gene. Die Vögel verhalten sich also so, dass sie eine möglichst hohe Gesamtfitness erreichen. Dies wird deutlich, wenn der Revierinhaber stirbt. Dann übernimmt ein männlicher Helfer seine Position. Also ist es keineswegs so, dass die Helfervögel auf das Brüten freiwillig verzichten. Sie machen nur das Beste aus dem Umstand, dass sie nicht brüten können. Sie erreichen somit eine höhere Gesamtfitness als Nichthelfer, wodurch sie selektiv im Vorteil sind.
2.2.2.2 Fallbeispiel Honigbiene
Die Honigbiene Apis mellifica ist ein besonderes Beispiel für die Anwendung der Theorie der Verwandtenselektion. Die Bienen stellen eine Besonderheit dar, weil nur eine weibliche Biene pro Staat fruchtbar ist und zwar die Königin. Alle anderen weiblichen Bienen, die auch als Arbeiterinnen bezeichnet werden, sind steril, haben aber wie die Königin auch einen diploiden Chromosomensatz. Die männlichen Bienen, die Drohnen, unterscheiden sich von den weiblichen Bienen dadurch, dass sie nur einen Chromosomensatz besitzen, also haploid sind. Aus einer durch eine Drohne befruchtete Eizelle wird eine Arbeiterin oder eine König und aus einer unbefruchteten Eizelle eine Drohne.
Nun verhält es sich so, dass die Arbeiterinnen der Königin bei der Eiabgabe helfen und praktisch die gesamte Aufzucht übernehmen

Wieder muss dieses Verhalten selektiv vorteilhaft sein. Um dies zu verdeutlichen, bedarf es einer kleinen Tabelle. Wir betrachten einen Fall in dem die Drohnen, die die Königin befruchtet haben, aus ihr hervorgegangen sind. Die Drohnen sind also Söhne der Königin. Die Königin habe die beiden Allele (A) und (B). Also haben die Drohnen entweder (A) oder (B). Es ergibt sich damit nach Mendel für Arbeiterinnen der nächsten Generation folgende Kombination mit den relativen Häufigkeiten: AA=25%, AB=50%, BB=25%.
Die Tabelle zeigt den Verwandtschaftsgrad (r) zwischen den Geschwistern aus diesen Beziehungen unter Berücksichtigung der relativen Häufigkeiten:
| |
AA |
AB |
AB |
BB |
| AA
| 1 |
0,5 |
0,5 |
0 |
| AB
| 0,5 |
1 |
1 |
0,5 |
| AB
| 0,5 |
1 |
1 |
0,5 |
| BB
| 0 |
0,5 |
0,5 |
1 |
Das ergibt einen mittleren Verwandtschaftsgrad r=0,625, der natürlich nur einen Durchschnittswert darstellt. Geht man davon aus, dass die Arbeiterinnen, die jetzt gerade die Eiablage unterstützen, ebenfalls auf diese Weise entstanden sind, so sind die befruchteten abgelegten Eier quasi ihre Schwestern, mit denen sie dann der Verwandtschaftsgrad r=0,625 verbindet. Würden die sterilen Arbeiterinnen doch Eier ablegen können, würde sie mit ihren Nachkommen nur der Verwandtschaftsgrad r=0,5 verbinden. Sie sind mit ihren Schwestern also im Durchschnitt näher verwandt als mit ihren Kindern. Deshalb verhalten sich die Arbeiterinnen so, als wenn sie dies wüssten und erhöhen auf diese Weise ihre Gesamtfitness über das Maß hinaus, welches eine eierlegende Arbeiterin erreichen würde. Sie tragen also durch die Pflege ihrer Schwestern mehr zur Verbreitung ihrer Gene bei, als dies durch eigene Nachkommen möglich wäre.
2.2.3 Eine Krankheit rettet Leben: Die Sichelzellanämie in Afrika
In Afrika sterben jährlich 600 Millionen Menschen an Malaria. Menschen, die Gene enthalten, die wirksam vor Malaria schützen, sind also selektiv im Vorteil. Solche Abwehrstrategien einer Infektion haben sich deshalb schon vor langer Zeit in Afrika ausgebreitet. Eine besonders weit verbreitete ist die sog. Sichelzellanämie, die wirksam auch vor den gefährlichsten Malariaerregern schützt. Leider ist das nur die eine Seite der Medaille, denn mit der Anämie sind auch gesundheitliche Probleme verbunden. Doch die Selektion wirkt in Richtung der Anämie, weil Menschen mit ihr eine höhere Fitness aufweisen als Menschen ohne sie.
2.2.3.1 Überblick Malaria
Die Malaria auch Sumpffieber genannt, ist eine fast ausschließlich in den Tropen vorkommende Infektionskrankheit, die durch bestimmte Mückenarten der Gattung Anopheles übertragen wird.
Überträgertiere sind ausschließlich die Weibchen, da nur sie blutsaugend sind. Sticht eine infizierte Mücke einen Menschen, so gelangen mit dem Speichel der Mücke die unreifen Sporenformen (Sporozoiten,Sichelkeime) in den Organismus. Dort reifen sie in der Leber heran. Nach dieser 1-6 wöchigen symptomfreien Phase (Inkubationszeit) werden die ausgereiften Malariaerreger (Schizonten) , die sich zuvor ungeschlechtlich in Merozoiten geteilt haben, ins Blut ausgeschwemmt. In den roten Blutkörperchen (Erythrozyten) entwickeln sie sich wiederum Schizonten (Blutplasmodien), die sich durch ungeschlechtliche Teilung zu jeweils 8-12 Merozoiten vermehren. Schließlich kommt es zur Zerstörung der roten Blutkörperchen und damit zum Fieberanfall (Abb.12).
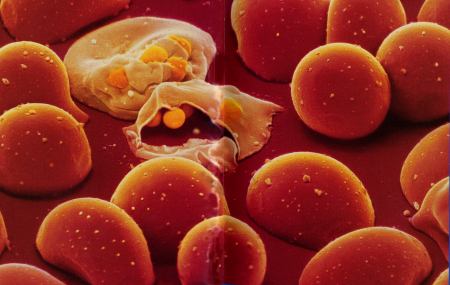
Dieser Vorgang wiederholt sich in bestimmten Zyklen, so dass zwischen ihnen keine Fieberschübe liegen. Nach 4-5 solcher Vermehrungszyklen entstehen in den Erythrozyten Geschlechtsformen (Gametozyten), die sich im menschlichen Körper nicht weiter vermehren können. Sticht nun eine nicht infizierte Mücken diese Person, so entwickeln sich in der Mücke wieder Sporozoiten, so dass sich der Kreislauf schließt. Diese Mücke wird zur potentiellen Gefahr für jeden Menschen.
2.2.3.2 Überblick Sichelzellanämie
Wie der Name schon sagt handelt es sich bei der Sichelzellanämie um eine Anämie, genauer gesagt um eine hämolytische Anämie, also um den verstärkten Abbau roter Blutkörperchen.
Die Sichelzellanämie ist genetisch bedingt. Die Vererbung erfolgt autosomal kodominant. Durch diesen genetischen Defekt kommt es zu einer Störung bei der Bildung des roten Blutfarbstoffes. Es wird abnormes Hämoglobin (sogenanntes Hämoglobin S) gebildet. Dieses hat im Vergleich zum normalen Hämoglobin eine viel geringere Löslichkeit, insbesondere unter Sauerstoffmangel im Blut. Dies führt zur Verklebung von Hämoglobinmolekülen mit Ausbildung starrer Formationen wodurch die roten Blutkörperchen die typische sichelförmige Gestalt annehmen (Abb. 13). Sie verlieren ihre Verformbarkeit, es kommt daher leicht zur Verstopfung kleinerer Blutgefäße.
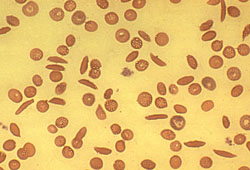
Menschen bei denen beide Allele Sichelzellallele sind (Homozygote), haben eine stark verminderte Lebenserwartung, da bei ihnen fast immer alle roten Blutkörperchen die Sichelform aufweisen.
Menschen mit einem Sichelzellallel und einem gesunden Allel (Heterozygote), haben normalerweise ganz normale rote Blutkörperchen. Bei starker Anstrengung, allgemein bei einem gesteigerten Sauerstoffverbrauch, kann es aber zur Umformung der roten Blutkörperchen kommen. Es kommt zum plötzlichen Zerfall einer großen Menge von roten Blutkörperchen. Diese Verkürzung der Lebensdauer der roten Blutkörperchen kann aber durch eine gesteigerte Produktion im Knochenmark ausgeglichen werden.
2.2.3.3 Sichelzellanämie und Malaria
Was hat das aber nun mit der Malaria zu tun? Nun, da die Merozoiten zur Fortpflanzung normale rote Blutkörperchen brauchen, gewährt die Sichelzellanämie einen gewissen Schutz gegenüber der Krankheit. Denn in Sichelzellen kann sich der Malariaerreger nicht vermehren und es kommt daher nicht zur Zerstörung roter Blutkörperchen.
Daher müssten homozygote Sichelzellallelträger einen starken Selektionsvorteil haben. Da sie aber durch die Sichelzellanämie eine stark verkürzte Lebensdauer haben, sind sie selektiv nicht im Vorteil und erreichen keine hohe Fitness.
Heteroygote Träger sind die selektiv erfolgreichste Gruppe. Ihre roten Blutkörperchen liegen zwar in normaler Form vor, aber hin und wieder kommt es zu einer sogenannten hämolytischen Krise, so dass sich die roten Blutkörperchen sichelförmig verformen. Aufgrund der Tatsache, dass sich der Körper daran gewöhnt hat, haben heterozygote eine erhöhte Produktion an roten Blutkörperchen, wodurch bei einer Malariaerkrankung schnell neue Blutkörperchen nachgebildet werden, so dass die Sterblichkeitsrate bei einer Erkrankung sinkt.
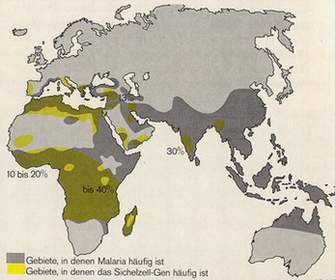
Deshalb sind Heterozygote in Malariagebieten selektiv im Vorteil. In nicht Malariagebieten sieht dies anders aus wie Abbildung 14 zeigt. Dort ist das Sichelzellgen nicht verbreitet. Nur in Malariagebieten und der afrikanisch stämmigen amerikanischen Bevölkerung erreicht es eine hohe Verbreitung.
3. Schlussbetrachtung
Hier sei dem Verfasser noch eine kleine Anmerkung erlaubt, die mit der eigentlichen Biologie, zwar nicht viel gemein hat, aber den Versuch macht, die biologischen Erkenntnisse im Bereich der Evolutionstheorie, speziell aber im Bereich der Selektion, historisch einzuordnen und zu bewerten.
3.1 Falschverstandene Selektion
Kann man das Selektionsprinzip aus Darwins Evolutionstheorie auch auf den Menschen anwenden? Diese Frage kam natürlich auf, nachdem Darwin mehr als 20 Jahre nach seiner Forschungsreise am 1.7.1858 seine Evolutionstheorie das erste Mal vortrug.
Eine pauschale Beantwortung dieser Frage ist sicher nicht möglich, doch muss man sich in die Zeit zurückversetzen. Europa war nach dem napoleonischen Zeitalter neu geordnet worden. Der Einfluss, den Europa auf seine Kolonien in Übersee auszuüben vermochte, wurde geringer. Doch die Industrie, die auf extraeuropäische Rohstoffquellen angewiesen war, fürchtete den Verlust. Es war 1882, als mit dem Anschluss Ägyptens an das britische Empire der Startschuss für den Wettlauf um Afrika fiel.
Ein ganz wesentlicher Faktor für diesen Schritt war die Idee des Sozialdarwinismus, welcher die oben erwähnte Frage klar mit Ja beantwortete. Diese pseudowissenschaftliche Theorie besagt, dass es wie in der Natur auch bei den Menschen darauf ankommt, der "Stärkste" zu sein, denn nur jener würde im Kampf der Kulturen und Nationen gewinnen. Ich verweise hier noch einmal auf die Kernaussagen Darwins zur Selektion, die unter 1.1 auf Seite 2 aufgeführt sind. Jede einzelne europäische Nation war wie besessen von dieser These. Wie mag die Einnahme Ägyptens durch England auf die anderen europäischen Mächte gewirkt haben? Nach der sozialdarwinistischen These müssten sie, um Schritt zu halten ebenfalls neue Landstriche in Besitz nehmen, damit England nicht "selektiv im Vorteil" bliebe. Der Nationalstolz nahm solche Dimensionen an, dass bis zum Anfang des 1. Weltkrieges ganz Afrika unter den europäischen Mächten aufgeteilt war, alles getarnt unter dem Deckmantel der Kulturbringermission, nach dem Motto:"Die armen Wilden müssen endlich lernen was Kultur ist. Sie haben ja keine."
Doch dies war noch lange nicht das schlimmste, wofür dieser, rein wissenschaftlich betrachtet, großartige Einblick in die Funktionsmechanismen der Natur missbraucht wurde. Der Nationalsozialismus überzog mit seiner Rassenlehre ganz Europa und vergiftete selbst fähigste Köpfe mit seinen wirren Ideen. In der sog. Rassenlehre, eine Bezeichnung die einen auf wissenschaftlichen Tatsachen beruhenden Hintergrund vermuten lassen soll, wurde behauptet, dass Menschenrassen unterscheidbar seien und dass diese Rassen einen unterschiedlichen Wert haben. So wurde der Arier als die Krönung der Schöpfung bezeichnet, während vor allem Juden, Sinti und Roma und Russen als Untermenschen bezeichnet wurden, die es nicht wert waren zu leben. Diese Völker würden nur wertvollen "Lebensraum" in Anspruch nehmen. Man beschleunigte nur die Selektion, wenn man sie sofort tötete. Diese Rassenlehre ist aber völlig aus der Luft gegriffen, da man gar nicht von Menschenrassen sprechen kann. Dazu ist es in Europa viel zu sehr zu Durchmischungen des Erbgutes gekommen. Es gibt keinen Arier oder eine deutsche Rasse, es gibt nur Menschen. Wahrscheinlich waren viele Juden "deutscher", als so mancher böhmische Gefreite1. Aber auch vor "Volksdeutschen" machte dieses System nicht halt. So wurden zehntausende behinderte Menschen getötet, weil sie das deutsche Blut verunreinigen würden.
Genau hier muss man sich fragen, was den Menschen vom Tier unterscheidet, wenn man einen Unterschied zulassen will, was aber in dieser Frage wohl zweifelsohne feststeht. Es herrscht eben kein Kampf ums Dasein. Es ist ein Miteinander in unserer Gesellschaft. Darauf muss man sich immer wieder besinnen.
Ich möchte diesen Abschnitt mit den Worten schließen, die die Schuldirektorin des Gutenberg-Gymnasiums in Erfurt unter die Schülerzeitung geschrieben hat: "Mensch bleib' menschlich!".